Members
Harukazu Nakamura (Developmental Neurobiology)

Prof. Nakamura graduated from Kyoto University in 1971. After graduation he had been working at the Department of Anatomy of Kyoto Prefectural University and Hiroshima University for 17 years. He got his phD in Hiroshima University. He then became a professor of Biology at Kyoto Prefectural University of Medicine, and moved to Institute of Development, Aging and Cancer, Tohoku University in 1994. He has participated in the Graduate School of Life Sciences from the first (2001). He learned micromanipulation of chick embryos from Prof. Nicole Le Douarin (1979-1980), and has been working on brain pattern formation in chick embryos since then. He established the system of gene transfer to living chick embryos by electroporation in ovo, which revived the chick embryo as a model animal for developmental biology, and this method is now a routine technique in the study of developmental biology. His team greatly contributed to understanding the mechanisms of brain pattern formation. He headed the 'Brain Pattern Formation' team of Grant-in-Aid for Scientific Research on Priority Areas- of the Ministry of Education, Culture, Sports, Science and Technology of Japan during 1999-2004. He has been a chief editor of Development, Growth & Differentiation, the official journal of Japanese Society of Developmental Biologists (JSDB). He organized Annual Meeting of JSDB in 2005. He published books sharing authorship, 'Neuroscience, editor Masao Ito, from Miwa Shoten'; and 'Standard Cell Biology, from Igaku Shoin'. He also translated textbooks into Japanese, 'Human Embryology of Larsen' and 'Physiologie du Neurone, eds Tritsch et al., Doin editeru-Paris'.
- Introduction of Research
-
We have been interested in the regionalization of the vertebrate central nervous system (CNS). We have developed a very effective gene transfer system in chick embryos, in ovo electroporation. This system is so effective for misexpression of the gene of interest that the chick embryo is revived as a model animal for developmental biology. The system is applied in other animals, and become routine technique in developmental biology. siRNA system has enabled us to silence the expression of the target gene, which could also be applied by electroporation.
We have studied the mechanisms of regionalization of the optic tectum and cerebellum. It is now accepted that the fate of the brain vesicles is determined by combination of the transcription factors expressed, and that organizing molecule emanated from the local organizing center regulates expression of the transcription factors to determine the fate of the adjacent region. Midbrain hindbrain junction (isthmus) functions as a local organizing center emanating Fgf8. We have shown that strong Fgf8 activates Ras-ERK signaling pathway to induce cerebellar differentiation. If this signaling pathway is disrupted by dominant negative form of Ras, the optic tectum differentiates in place of cerebellum. But this signaling pathway should be regulated precisely for mesencephalon metencephalon boundary formation. If it overflows the boundary shifts anteriorly, and if the signaling is repressed the boundary shifts posteriorly. We have also shown that the region where Otx2, Enl and Pax2 are expressed differentiates as mesencephalon. Repressive interaction between Pax6 and En1/Pax2, and between Otx2 and Gbx2 determines the di-mesencephalic and mes/metencephalic boundary, respectively. We are now interested in downstream signal transduction of Fgf8 in the midbrain and hindbrain. We also pay attention to the polarity formation in the tectum.
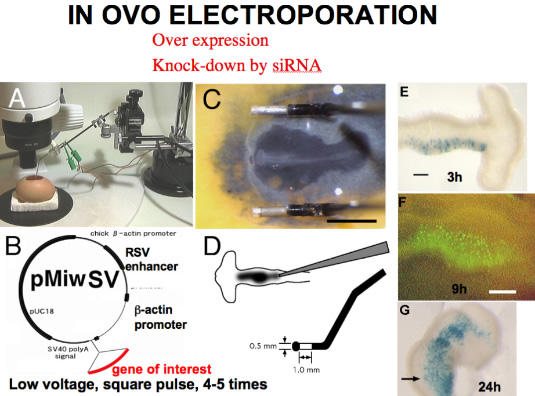
Chick embryos had been used as a model animal of experimental embryology, but difficulty in manipulating the embryo while in the uterus interfered with molecular approach to the embryo (such as production of transgenic chick or of gene-knock-out chick). Development of gene transfer by electroporation in ovo in our lab revived chick embryos as a model animal of modern developmental biology. Combination of siRNA and electroporation made it possible to knock-down a targeted gene expression. Moreover, we can now integrate transgene by transposon system, and follow long term effects of the tranfered gene. Now we can carry out gain and loss of function of a gene of interest in chick embryos at desired place and stage of the chick embryo.
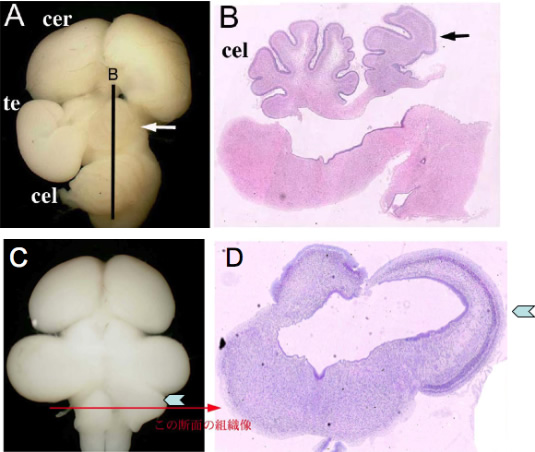
Fate change of the mesencephalon to the cerebellum by Fgf8b (A, B), and reverse by dominant negative form of Ras (C, D). These figures suggest that strong Fgf8 signal activates Ras-ERK signaling pathway to differentiate a cerebellum.
- Articles
-
- Suzuki-Hirano, A., Sato, and Nakamura, H. (2005) Regulation of isthmic Fgf8 signal by Sprouty2. Development, 132, 257-265.
- Sato, T. and H. Nakamura (2004) The Fgf8 signal causes cerebellar differentiation by activating Ras-ERK signaling pathway. Development 131, 4275-4285.
- Sato, T., Araki, I., Nakamura, H. (2001) Inductive signal and tissue responsiveness to define the tectum and the cerebellum. Development, 128: 2461-2469.
- Katahira, T., Sato, T., Sugiyama, S., Okafuji, T., Araki, I., Funahashi J., and Nakamura, H. (2000) Interaction between Otx2 and Gbx2 defines the organizing center for the optic tectum. Mech. Dev. 91, 43-52.
- Nakamura, H. (2001) Regionalisation of the optic tectum: Combination of the gene expression that defines the tectum. Trends Neurosci. 24, 32-39.